1. L'origen de la Terra




Actualment la major part de la comunitat científica accepta la teoria del Big Bang per explicar l'origen de l'Univers: fa uns 15.000 milions d'anys tota la matèria i l'energia estaven concentrades en un punt (teoria de l'ou còsmic), i a causa de la inestabilitat gravitatòria i d'altíssima temperatura, es va produir la Gran Explosió. En els instants posteriors es van començar a formar les primeres partícules subatòmiques, que van donar origen als àtoms alhora que disminuïen la temperatura i la densitat de l'Univers, que encara avui està en expansió.
El sistema solar es va originar fa uns 5000 milions d'anys a partir d'una nebulosa és a dir un núvol còsmic format per gasos i pols interestelar (molècules de diferent grandària, de 0,1 a 100 micres, essent majoritàriament gels i roca, és a dir, cadenes de silici). La matèria es va començar a concentrar en un extrem de la Via Làctia on es va iniciar la contracció gravitatòria. La major part de la massa es va situar al centre de la nebulosa, i l'enorme temperatura assolida va permetre l'inici dels processos de fusió nuclear. El nucli molt més dens es va convertir en una estrella i va començar a brillar; així va néixer el Sol.
Les restes de núvol també es van condensar, es van agrupar independentment i van formar cossos cada vegada més grans, primer planetesimals després protoplanetes, que finalment van acabar sent els planetes i satèl·lits actuals, entre els quals es troba la Terra.
1.1. Formació de la Terra
L'aspecte actual de la terra és molt diferent del que tenia originalment. En el període inicial de la seva formació, era una massa incandescent i, a mesura que el protoplaneta creixia, s'anava escalfant. L’increment de temperatura era causat, majoritàriament, per la contracció gravitatòria i per l’acreció* de la matèria així com per l'impacte dels meteorits i la desintegració radioactiva dels isòtops com l’urani (U) i el tori (Th). Una part d'aquesta calor primitiva encara es conserva al nucli de la Terra.
Està estructurada en capes, segons la seva composició i densitat. Quan el protoplaneta va assolir la mida definitiva la matèria fosa va començar a emetre calor a l'espai per radiació. La temperatura va anar disminuint i el planeta es va anar refredant lentament. Les capes més externes van ser les primeres que es van solidificar .
Amb la davallada tèrmica es va produir una diferenciació geoquímica dels materials:
a la geosfera (part “sòlida” o de roca de la Terra), els elements més densos es van enfonsar cap a l'interior terrestre i van formar el nucli, Els més lleugers es van quedar al superfície i van constituir l’escorça i el mantell primitius.
La hidrosfera (part “líquida” i formada per aigua), menys densa, es va situar sobre la superfície de la geosfera.
encara amb menor densitat i per tant sobre les dues anteriors es va formar l'atmosfera (part gasosa del planeta).
*acreció: formació de masses grans, com ara planetes i satèl·lits o cometes, com a resultat del xoc i l'adhesió entre si de partícules petites de pols i gels de diversa composició.
1.2. Formació dels oceans i l'atmosfera
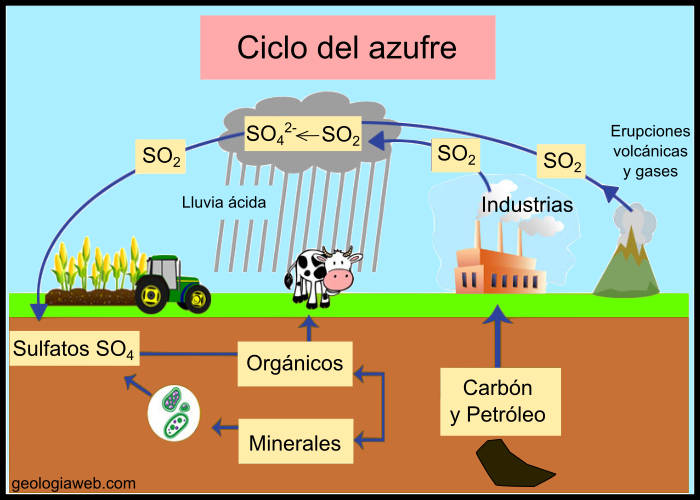
Al principi, la Terra no tenia atmosfera i els meteorits impactaven constantment contra la seva superfície, la qual cosa provocava un augment de la temperatura. L'activitat volcànica era molt intensa I s’emetien una gran quantitat de gasos a l'exterior. Aquests gasos, juntament amb els procedents del desgasament de l'interior del planeta i les substàncies volàtils de les primeres fases de formació, Van configurar la Primera capa de la Terra l'atmosfera primigènia.
Aquesta atmosfera inicial contenia enormes quantitats de diòxid de carboni i vapor d'aigua. Quan la temperatura va baixar i la Terra primitiva es va anar refredant, es van condensar grans quantitats d'aigua i es van produir les primeres pluges, que van començar a omplir les depressions i van formar mars i oceans. El naixement de la hidrosfera és un punt d'inflexió en l'evolució del nostre planeta, ja que el seu origen va seguit de l'aparició de la vida.

2. Datació i mètodes de datació
En geologia és molt important datar els diferents esdeveniments que han anat tenint lloc per poder reconstruir la història geològica i ordenar-la cronològicament. La manera més eficaç d'aconseguir-ho és estudiar amb profunditat les roques i els fòssils. Hi ha dos tipus de datació:
- La datació absoluta indica l'edat exacta de l'element (ja sigui una roca o un fòssil), és a dir, dóna un valor numèric concret, una edat real en milions d'anys.
- La datació relativa ofereix la possibilitat d'ordenar els successos del més antic al més nou, de comparar els esdeveniments, però sense atribuir-los una edat concreta.
2.1. L'edat de les roques: mètodes de datació
No totes les roques aporten la mateixa informació, ni en totes es poden utilitzar els mateixos mètodes de datació: el mètode variarà segons la composició i l'origen de la roca.
DATACIÓ ABSOLUTA: MÈTODE RADIOMÈTRIC
És una datació mitjançant isòtops radioactius (tipus d'àtoms del mateix element químic amb diferent nombre de neutrons); es basa en la presència d'elements radioactius en els minerals que componen la roca.
Els àtoms dels isòtops radioactius són inestables i es desintegren per transformar-se en elements més estables. Durant aquest procés alliberen energia, la radiació, i la velocitat a la qual ho fan és constant i es pot conèixer amb exactitud. Cada isòtop té un període de semidesintegració o semivida concret, essent el temps necessari perquè es desintegrin la meitat dels àtoms radioactius de la mostra.
Aquest temps varia molt segons el tipus d'àtom del qual es tracti, des de dies fins a milions d'anys. Per exemple:
- l'urani-235 es transforma en plom-207 i té una semivida de 704 milions d'anys
- el iode-131 passa a xenó-131 i té un període de semidesintegració de 8,02 dies.
Això permet establir uns autèntics rellotges de datació absoluta. El següent gràfic mostra la corba de desintegració del carboni-14 en nitrogen-14. A l'eix vertical es representa la quantitat d'àtoms de l'element radioactiu inicial, en %, i a l'eix horitzontal el temps que transcorre des de l'inici de la desintegració. La massa del 14C es redueix a la meitat cada 5370 anys, i l'altra meitat es desintegra espontàniament en 14N.
No obstant això, aquest mètode únicament és vàlid per datar les roques ígnies, les que es formen a partir d'un magma: en solidificar-se contenen el 100% dels isòtops radioactius, que comencen a desintegrar-se. Per conèixer l'edat de la roca, es mesura la quantitat d'àtoms radioactius i d'àtoms estables que conté, i es calcula el temps que fa que es va formar.
En l'exemple anterior, el de la datació a partir del carboni-14, es tracta d'una de les tècniques més fiables per conèixer l'edat de mostres orgàniques. S'utilitza per fer datacions molt recents (des del punt de vista geològic); fins fa poc, no passava dels 10000 anys, però actualment les noves tècniques permeten arribar als 60000. S'aplica, sobretot, en arqueologia, antropologia i geologia del Quaternari (la darrera era geològica, la més moderna).
És una pràctica basada en la disminució exponencial d'aquests isòtops radioactius., amb 6 protons i 8 neutrons al nucli i que trobem a tots els organismes. El carboni prové, com ja s'ha vist en la unitat anterior, del diòxid de carboni de l'atmosfera; alguns isòtops són 14C, tot i que hi són presents en una proporció menor que el 12C (1 carboni-14 per cada bilió de carboni-12); no és l'únic isòtop del carboni, n'existeixen dos més (el 11 i el 13). En el procés de la fotosíntesi les plantes incorporen tots els tipus de carboni des del CO2, i els animals també a través de les cadenes tròfiques. Quan un ésser viu mor, comença la desintegració del carboni-14 i la transformació en nitrogen-14, i quan n'han transcorregut 5730 anys el valor de la seva semivida s'haurà reduït a la meitat.
Un altre exemple:
L'urani-238 és un isòtop radioactiu que es descompon i es transforma en un altre element més estable, el plpom-206. El període de semidesintegració del 14U és de 4470 Ma (milions d'anys), pràcticament l'edat de la Terra. Aquest fet significa que, quan han passat 4470 Ma, la meitat de la massa d'urani s'ha desintegrat i ha format plom, i així successivament.
DATACIÓ RELATIVA: MÈTODES ESTRATIGRÀFICS
Com que la tècnica radiomètrica no és vàlida per a les roques sedimentàries, cal recórrer a altres mètodes de datació relativa, com els estratigràfics.
L'estratigrafia és la disciplina de la geologia que estudia i interpreta les roques sedimentàries i els estrats o capes que les formen.
Per poder ordenar els diferents esdeveniments i establir una escala de temps relatiu, cal aplicar els tres principis bàsics de l'estratigrafia:
- Principi de la superposició d'estrats (Niels Stensen -Nicolaus Steno-, 1669): els estrats superiors són més moderns que els inferiors, sempre que la seqüència de roques sedimentàries no estigui deformada. Per tant, segueixen l'ordre en el qual es van dipositar els materials.
- Principi de successió biològica (William Smith, 1816-1819): els fòssils d'una roca són contemporanis a la seva formació i, per tant, tenen la mateixa edat. Si trobem els mateixos organismes fòssils en dos estrats podem deduir que tots dos es van formar durant la mateixa època.
- Principi de l'actualisme o uniformisme (James Hutton, 1788): els processos naturals que han actuat en el passat són els mateixos que actuen en el present. Per tant, si estudiem el present, podrem deduir i interpretar el passat i, en certa manera, predir el futur.